
| TEAM |
In the subsequent text we show some results obtained within the SP 5: Acquisition of productive, ecological, environmental and sociological data in forest environment. The results are grouped in two thematic focuses: A.) aerial studies implementing UAS Multispectral Imaging and B.) terrestrial studies based on direct tree measurements and destructive sampling. The results shown here are just brief selection from the obtained findings and would be considered as preliminary (“half-time”) outputs.
A. Tree Species Classification and Health Status Assessment for a Mixed Broadleaf-Conifer Forest with UAS Multispectral Imaging
Automatic discrimination of tree species and identification of physiological stress imposed on forest trees by biotic factors from unmanned aerial systems (UAS) offers substantial advantages in forest management practices. Given the fact that biotic agents have a continuous negative impact on forest resources, affecting several species and forests globally, this study provides a new framework about the applicability of novel remote sensing methodological approaches, through complex statistical analysis for monitoring-mapping tree health status, aiming to support precision forest management and decision-making for small-scale areas (Fig. 1). In this context, the main objectives were to test how bi-temporal aerial multispectral (5-band) imagery data can be utilized for:
1) Testing the sensitivity of specific spectral indices for classifying tree species.
2) Health status assessment by assessing and discriminating crown symptoms.
3) Detailed tree species classification and health status assessment by support vector machine (SVM) algorithm.

Fig. 1. Map perspective of the actual study area in Kostelec. (a) Shows the captured ortho-mosaic (RGB) of the study area by UAS at spring and (b) shows the captured ortho-mosaic (RGB) of the study area by UAS at summer.
- Spectral analysis
Photogrammetric processed datasets achieved by the use of structure-from-motion (SfM) imaging technique, where resulting digital terrain models (DTMs), digital surface models (DSMs), and orthophotos with a resolution of 0.05 m were utilized as input for canopy spectral analysis, as well as texture analysis (TA). To develop a deeper understanding of the spatial and temporal changes in the pattern (Fig. 2) caused by seasonal changes (spring and summer 2019), several vegetation indices (VIs) including a variety of wavelength bands (RGB, red-edge, and NIR) were applied to evaluate the amount of vegetation cover change between the two seasons (Fig. 3 and Table 1).

Fig. 2. Examples of zoomed tree canopies on different tree species and health status from both seasonal periods in RGB spectral bands.
Table 1. Reflectance indices used in chronological order.
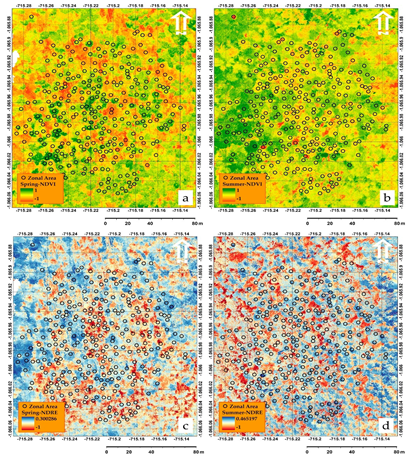
Fig. 3. (a) Showing the application of normalized difference vegetation index (NDVI) in spring (b) NDVI in summer (c) showing the normalized difference red-edge index (NDRE) in spring and (d) NDRE in summer.
From the post-hoc analysis (Fig. 4) NDVI, SAVI, and MCARI2 performed the best in classifying the reference data from spring 2019, whereas SAVI, MCARI2, and PSRI had the best performance for the summer period (Fig. 5). The above-mentioned VIs were able to successfully distinguish all the major classes from each other. Since the mentioned VIs are based on NIR, red, and red-edge wavelengths, they can detect the changes occurring in the amount of chlorophyll absorption and reflectance of the cellular structure of the leaves.
Furthermore, the post-hoc results highlighted an issue for the differentiation between major classes in spring between healthy broadleaves (B) and dead conifers (D), which likely occurred because of the nature of broadleaves being without leaves during the spring. This issue resulted in confusion in the identification of major classes in the case of RESR, M3CI, CI, PSRI, and NDRE VI layers.
According to the coefficient of determination Eta-squared (?2), MCARI2 and CI respectively had the highest correlation with field data major classes with Eta-squared (?2) of 0.73 in spring 2019 and 0.61 for summer 2019. These results confirms the importance of the red-edge band for detail classification of both tree species type and health status during the vegetation period, due to the sensitivity of the red-edge band to small variations in the chlorophyll content and consistent across most species and health status classes.
Post-hoc results for the summer exhibited better performance of PSRI compared to NDVI in the classification of reference data in different major classes. The results of Eta-squared (?2) were used to select the best independent variables, including VI and TA, for further analysis with the SVM.

Fig. 4. Post-hoc analysis for vegetation indices (VIs) showing the strength of association between the four major categorical classes (healthy broadleaves = B, healthy conifers = C, infested conifers = I,dead conifers = D) and the most significant descriptive statistics for each VI. Small letters (a,b,c,d) were used to denote significant differences between groups (p-value < 0.05).
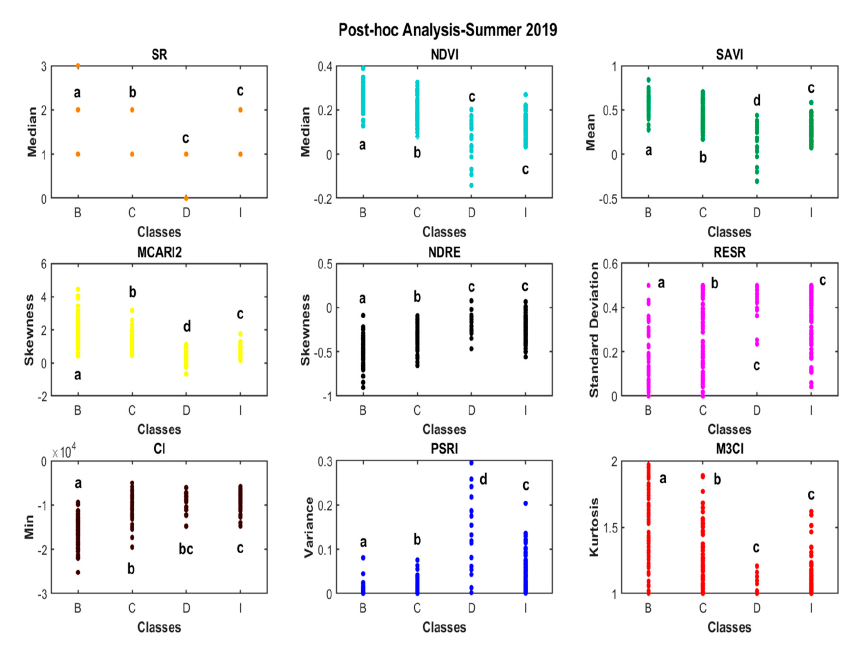
Fig. 5. Post-hoc analyses for vegetation indices (VIs) showing the strength of association between the four major categorical classes (healthy broadleaves = B, healthy conifers = C, infested conifers = I,dead conifers = D) and the most significant descriptive statistics for each VI. Small letters (a–d) were used to denote significant differences between groups (p-value < 0.05).
- Texture analysis
We map the TA within the zonal areas from topographic information (nCHM). For each season, 13 nCHM-based TA layers were mapped, including Mean, Variance, Entropy, Contrast, Heterogeneity, Homogeneity, Angular Second Moment, Correlation, Gray-level Difference Vector (GLDV) Angular Second Moment, GLDV Entropy, GLDV Mean, GLDV Contrast, and Inverse Difference, with a kernel window size of 25 × 25 (32-bit) (Fig. 6).
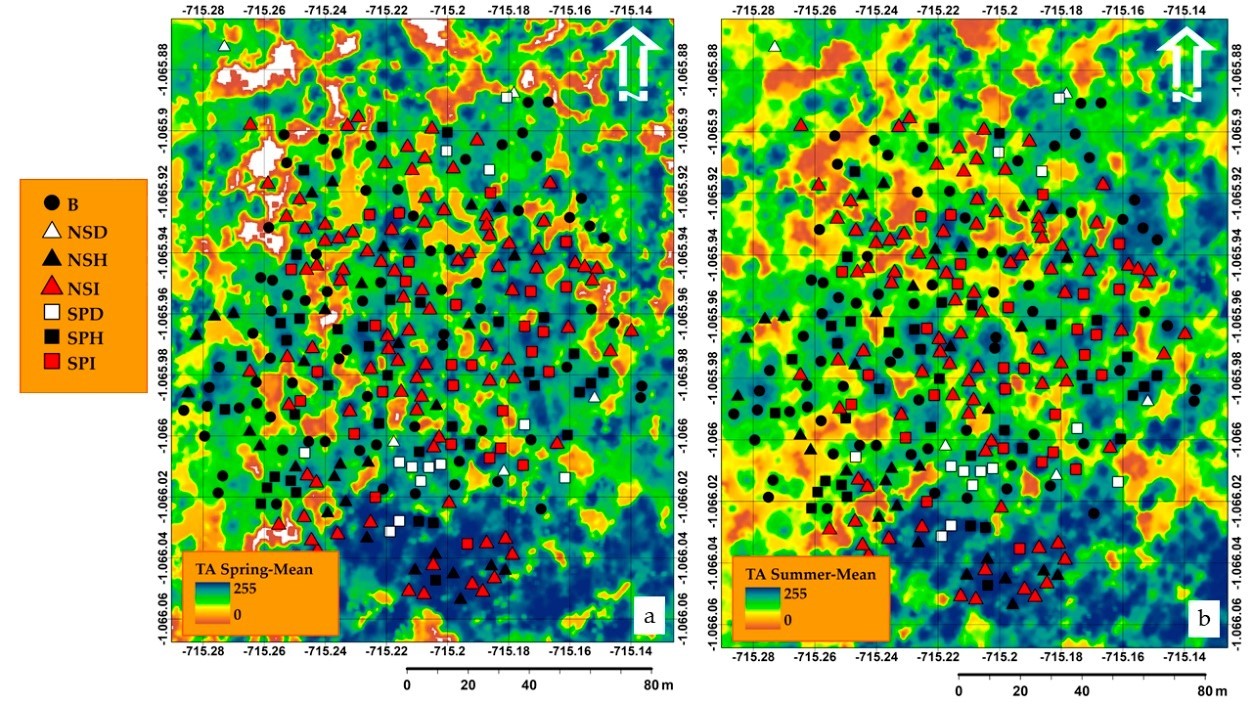
Fig. 6. Mean texture measure parameter applied in nCHM (a) in spring and (b) in summer, including the locations of healthy, infested, and dead trees based on their species classes (B = Broadleaves, NSD = Norway spruce dead, NSH = Norway spruce healthy, NSI = Norway spruce infested, SPD = Scots pine dead, SPH = Scots pine healthy, SPI = Scots pine infested).
Eta-squared (?2) exhibited that in most cases there was a weak or moderate relationship between predicted and observed values (e.g., Dissimilarity = 0.14, GLDV Angular Second Moment = 0.22, GLDV Entropy = 0.22, GLDV Mean = 0.14, Inverse Difference = 0.15, and Standard Deviation = 0.17), nCHM-based Mean was the only variable that showed a stronger relationship with the reference data compared to the rest of variables (Fig. 7), with an Eta-squared (?2) of 0.51 in spring 2019 and 0.28 for summer 2019.
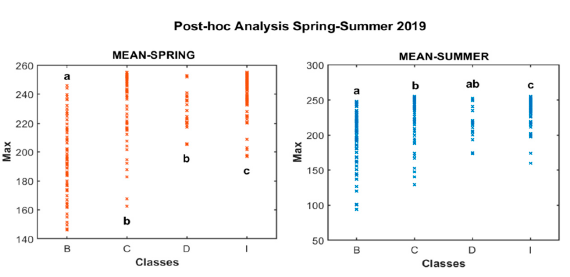
Fig 7. Post-hoc analysis for texture analysis (TA) showing the strength of association between the four major categorical classes (healthy broadleaves = B, healthy conifers = C, infested conifers = I, dead conifers = D) and the most significant descriptive statistic derived for the Mean variable in both seasons. Small letters (a–d) were used to denote significant differences between the groups (p–value < 0.05).
- Detailed tree species classification and health status assessment by SVM
In addition to the spectral and texture analysis that was applied for the general classification, the SVM algorithm was adopted for our unbalanced sample data to conduct a detailed classification of tree species and health status. For improving the accuracy of the output, the SVM classifier type 1 was used. The training and validation samples were randomly selected as 70% for training and 30% for validation of the modeling in each category. The SVM analysis showed significant performance for dividing the reference data into more complex classes that contained tree species and health status simultaneously.
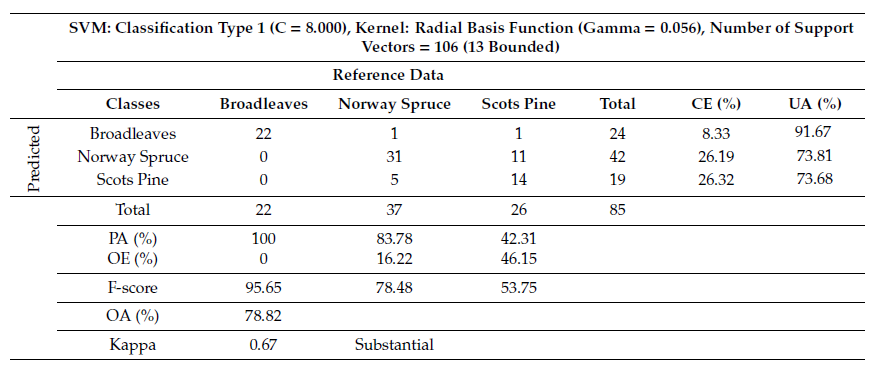
Based on Eta-squared (?2) results, all the VI layers which had strong relation with the reference data were used for modeling process using SVM. Those layers are: CI (Min) summer, CI (Median) spring; M3CI (Sum) spring, M3CI (Kurtosis) summer; MCARI (Mean) spring, MCARI (Skewness) summer; NDRE (Maximum) spring, NDRE (Skewness) summer; NDVI (Mean) spring, NDVI (Median) summer; PSRE (STD) spring, PSRE (Variance) summer; RESR (Variance) spring, RESR (STD) summer; SR (Mean) spring, SR (Median) summer; SAVI (Mean) spring, SAVI (Mean) summer. Modeling tree species using the above layers successfully distinguished the reference data into detailed classes, producing an F–score of 95.65% for broadleaves, 78.48% for Norway spruce and 53.75% for Scots pine,resulting in OA of 78.82% (Table 2).
To improve the model accuracy, fusion of the TA and VI layers was used for modeling the reference data. The criteria for the selection of TA layers for SVM were exclusively based on Eta-squared (?2) results. For TA layers we chose layers with moderate and strong relationship with the reference data, such as Mean (Mean) spring/summer; STD (Range) summer; GLDV Angular Second Moment (Skewness) summer; GLDV Entropy (Range) summer; GLDV Angular Second Moment (Coefficient of Variation) summer.
Table 2. Tree species classification summary table and confusion matrix using selected VI layers.
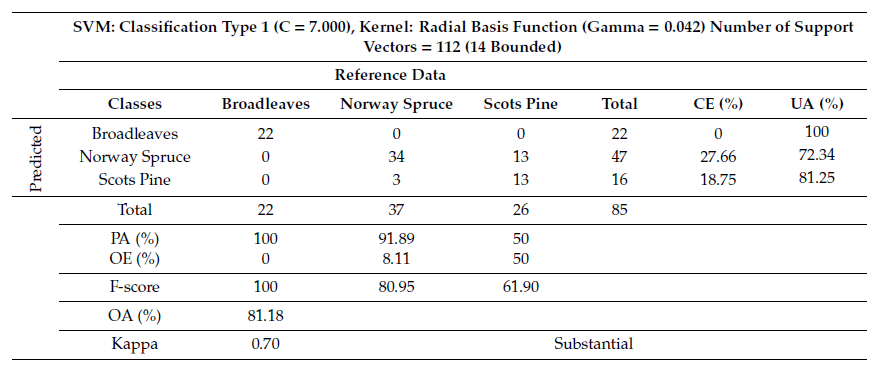
The combination of VI and TA layers successfully improved accuracy of modeling tree species using SVM by 2.36% (OA) (Table 3). SVM was able to separate broadleaves from the rest of the classes with an accuracy of 100%, resulting in both CE and OE of 0%. Similarly, SVM distinguish the Norway spruce from broadleaves and Scots pine with PA of 91.89%, resulting in an OE of 8.11% (Table 3). Consequently, the OA for distinguishing reference data for tree species from SVM was 81.18% (Table 3). We estimated a Kappa of 0.70, which suggested a strong (substantial) relationship between the reference data and the independent variables. In addition, the combination of selected layers improved the amount of F-score values of 100% for broadleaves, 80.95% for Norway spruce, and 61.90% for Scots pine.
Table 3. Tree species classification summary table and confusion matrix derived from the support vector machine (SVM) analysis using a combination of VI and TA layers.
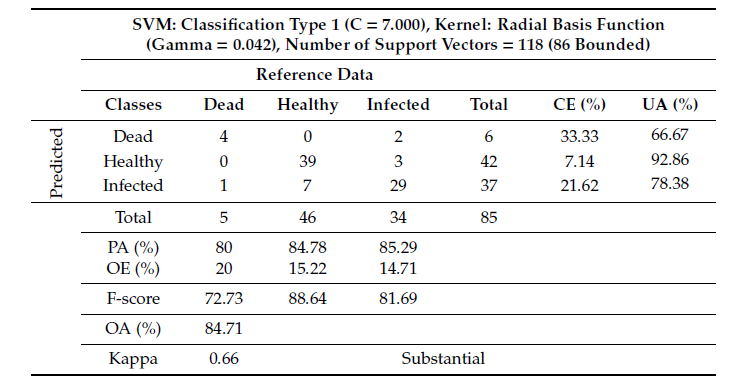
Considering only tree health status, combination of TA and VI layers could not improve the accuracy of health status classification. However, using solely VI layers the SVM algorithm distinguished the classes with a very high OA of 84.71% (Table 4). The PA for all tree health classes was found to be ?80%. Similar to the tree species classification, the Kappa for the assessment of health status was 0.66, which also suggested a substantial relationship between the reference data and the independent variables (Table 4).
Table 4. Classification table and confusion matrix of tree health status from the SVM using VI layers.
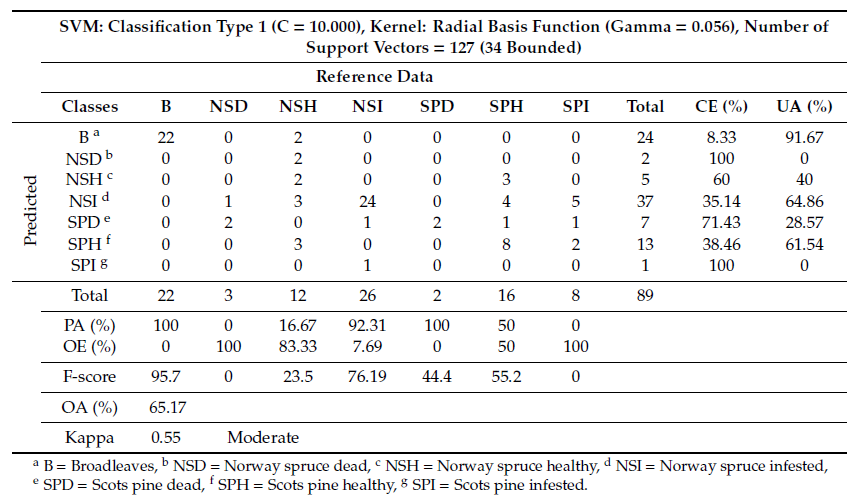
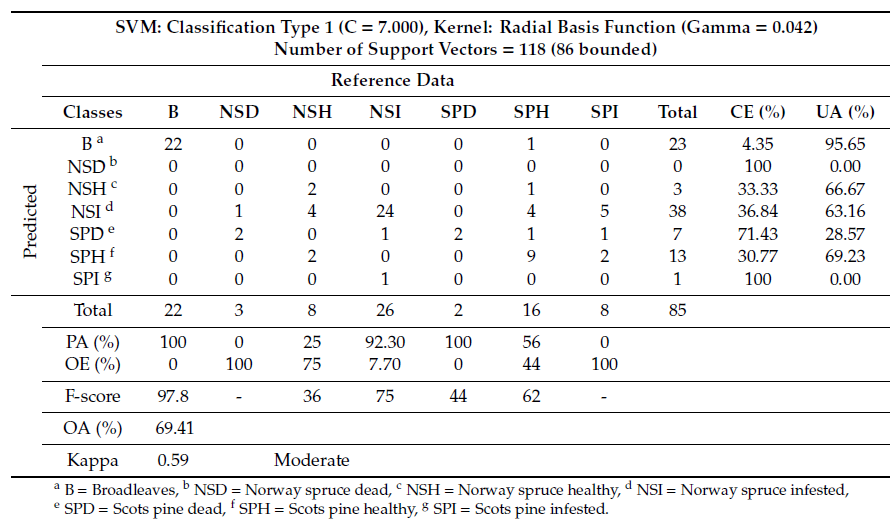
Classification of tree species/health using the selected VI layers successfully distinguished thereference data into detailed classes, producing an OA of 65.17% (Table 5).
Table 5. Classification summary table and confusion matrix including combined tree species and health status derived from the SVM analysis using VI layers.
Combination of TA and VI layers showed an increase in OA by 4.24% (Table 6). NSI and SPD classes were successfully distinguished with very high PA, 92% and 100%, respectively, which resulted in an OA of 69.41%. SPH yielded poorer, yet comparable, performance when compared to the NSI and SPD. When combining tree species and health status, the Kappa was poorer yet comparable (0.59), which represented a moderate relationship between reference data and independent variables (Table 6). Combination between texture and VI, improve significantly the distinguishment of coniferous species between healthy trees. The amount of F-score increased by 12.5% for healthy Norway spruce and 6.8% for the healthy Scots pine.
Table 6. Classification summary table and confusion matrix including combined tree species and health status derived from the SVM analysis using a combination of VI and TA layers.
In conclusion, due to the ability of UAS to acquire data in ultra-high spatial resolution, our method not only enabled mapping of the tree species and its current health state, but it also aids in monitoring the spread of disease, designating the possibility of either early-stage detection or new disease outbreaks to adjacent healthy trees. Furthermore, this method could be used to quickly evaluate areas that have been impacted by biological disturbance agents, for detection and mapping, tree inventory, and evaluating habitat conditions at relatively low costs. Conclusively, this approach can provide a new dimension of information derived from UAS platforms, which can lead to a better understanding of health status and tree species composition with reliable accuracy.
B. Terrestrial Measurements Related to Factors Influencing Tree (Stand) Properties
Aesthetic value (i.e. human perception of aesthetic features) relates especially to tree species composition, vertical and horizontal structure of stand. However, it is created also by individual esthetic “value” of each single tree. Esthetic value of individual tree is most probably linked to biomass allocation of aboveground biomass as well as morphological properties of foliage, branches and stem.
We expect that these properties on a tree level as well as stand level can be influenced for instance by:
1) Tree species as specific habit properties (aboveground biomass allocation into tree components).
2) Forest management, especially by means of stand density (crown competition) regulation.
3) Harmful agents, e.g. browsing of ungulate ruminating game.
In all three focuses we implemented terrestrial measurements and sampling on aboveground tree components. We are showing here just three examples of our aboveground biomass studies in the context of aesthetic properties of trees and stands regarding specific internal and external factors.
- Tree species as specific habit properties (aboveground biomass allocation into tree components)
We focused on measurements and modeling of aboveground biomass of eleven tree species with stem base diameter up to 100 mm. To determine quantity of the tree compartments, destructive method was used. Components were oven-dried and weighed (constant dry mass). Then, models, i.e. allometric relations for aboveground biomass were constructed by using stem base diameter as a predictor. The results have shown both quantitative (Fig. 8) and qualitative (biomass allocation) inter-specific differences (Figs. 9 and 10).

Fig. 8. Aboveground biomass models for some forest tree species.
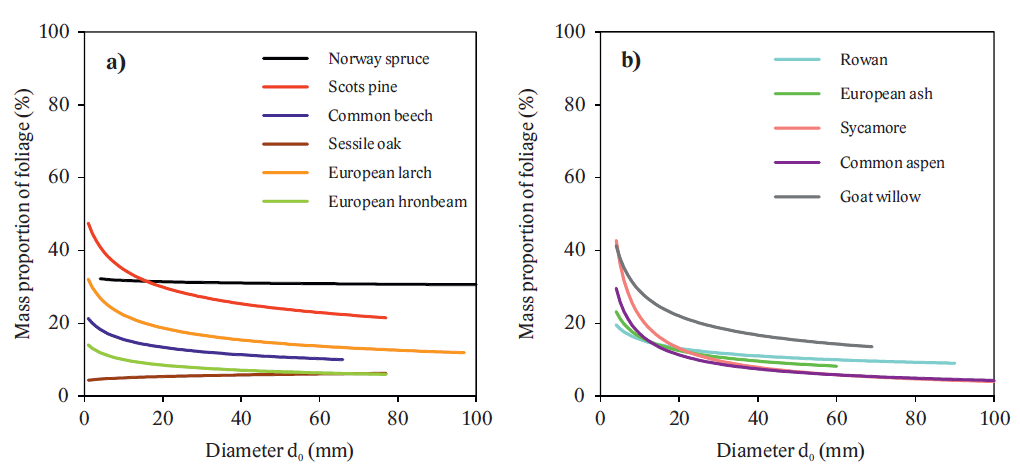
Fig. 9. Mass proportion of foliage to the aboveground biomass for the tree species
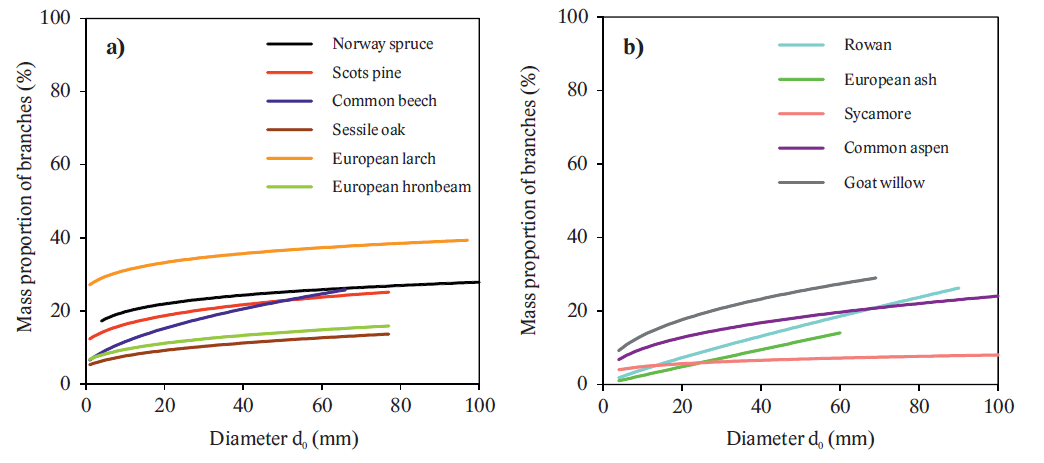
Fig. 10. Mass proportion of branches to the aboveground biomass for the tree species.
- Forest management, especially by means of stand density (crown competition) regulation
We researched aboveground biomass allocation in silver birch (Fig. 11) by means of destructive method. Birches under different crown competition were compared (classes: 0 - competition free crowns, 1 - mild competition, 2 - medium competition, and 3 - severe competition; see also Fig. 12).

Fig. 11. The birch-dominant stand studied for crown competition effects to biomass allocation (early spring season).

Fig. 12. Examples of the competition classes in sampled silver birches (bird view - dark colour indicates a target tree) recorded in the research site, specifically: 0 – competition free crown (scheme A), 1 – crown under mild competition (scheme B), 2 – crown under moderate competition (scheme C) and 3 – crown under severe competition (scheme D). The position of the trees and their crown projection were measured by the Filed-Map technology.
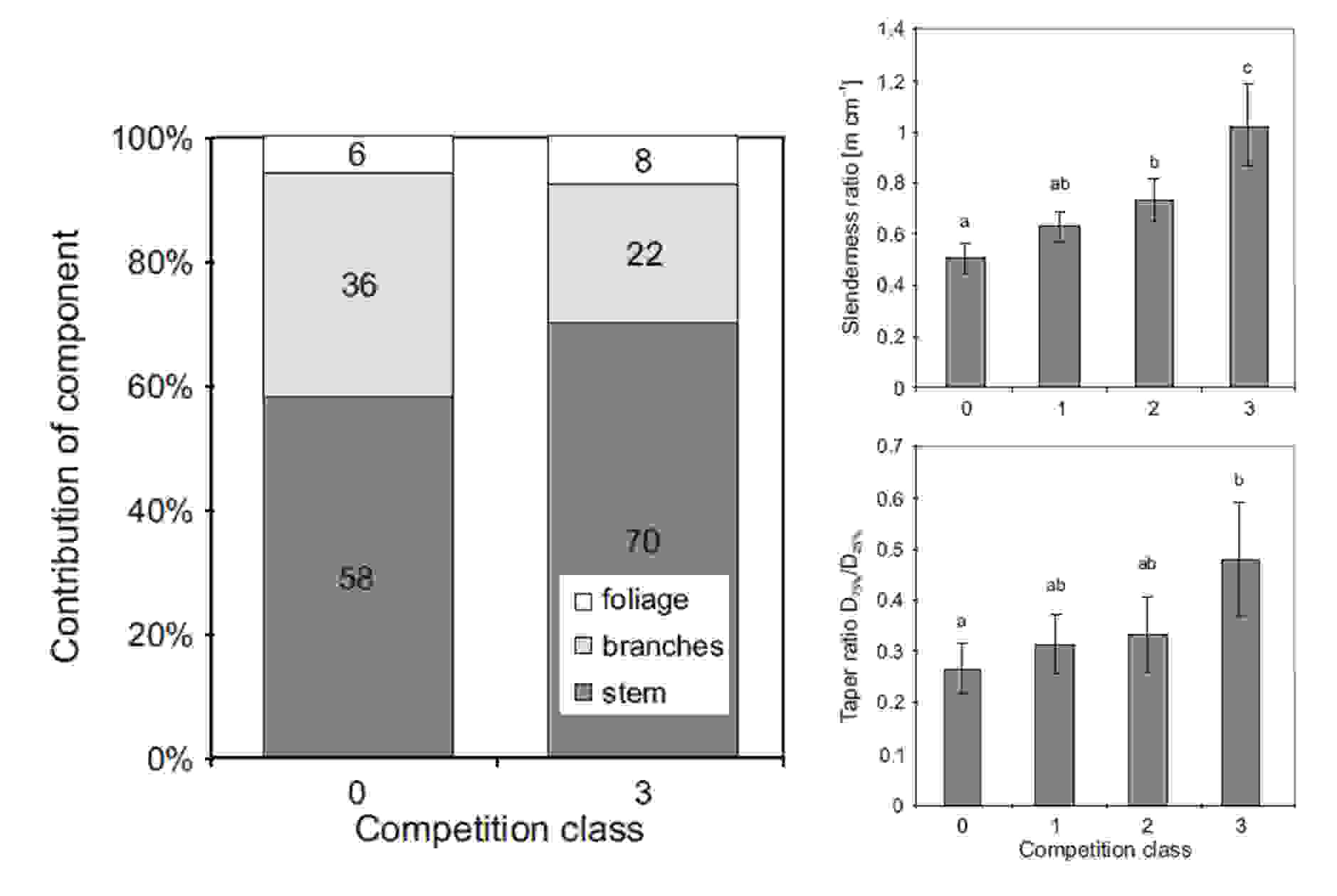
Fig. 13. Different biomass allocation between contrasting crown competition classes and stem shape characteristics in all crown competition classes.
The results indicate that stand density (possibly forest management regime), and consequently crown competition intensity influence biomass allocation and stem shape (Fig. 13). Less intensive competition stress means higher share of branches and more conic shape of stem.
- Influence of long-term browsing of ungulate ruminating game
We studied influence of ungulate ruminating game forage press to status of forest (production, species diversity, and spatial tree distribution). We established 15 circle-like plots inside fenced area (game-free) and 15 plots outside fenced area (under long-term browsing; Figure 14).
Fig. 14. Plots outside (purple circles) and inside (green circles) fenced area for quantification of browsing effects to forest.
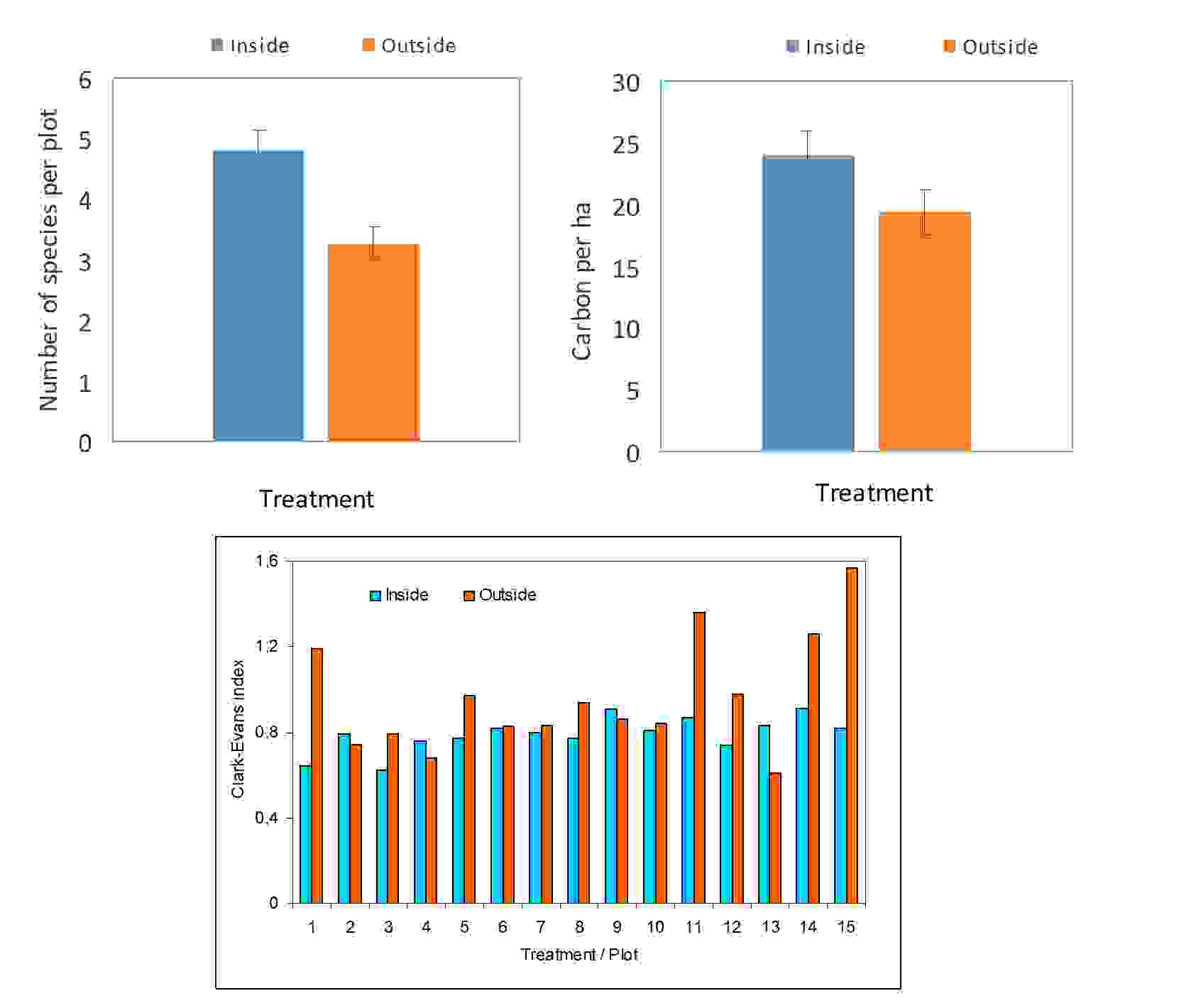
Fig. 15. Comparison of number of species per plot and carbon in aboveground biomass (t per ha) – upper diagrams, and values of Clark-Evans index for individual research plots – lower diagram.
The measurements showed that long-term game browsing decreased species diversity and aboveground biomass (carbon in tons per ha) stock (Fig. 15. upper diagrams). On the other hand - rather surprisingly, trees grown outside the fenced area had more regular spatial distribution (mean value of Clark-Evans index was 0.96) than trees grown inside the fenced area (0.79; Fig. 15. lower plate). All three experiments presented here indicates that qualitative and quantitative properties of individual trees as well forest stand as a whole are much modified by internal, i.e. species-specific and external, i.e. intentional management or natural conditions, factors.
Since some additional measurements and analyses are still needed, thus, more exact interpretation of these relationships, especially towards aesthetic value and its human perception will be prepared in the final stage of the project course.